| HOME | HELP | FEEDBACK | SUBSCRIPTIONS | ARCHIVE | SEARCH | TABLE OF CONTENTS |
| Cancer Research | Clinical Cancer Research |
| Cancer Epidemiology Biomarkers & Prevention | Molecular Cancer Therapeutics |
| Molecular Cancer Research | Cell Growth & Differentiation |
Articles |
Departments of Oncology Research [J. F. R., S. H., C. D., B. G., P. G-C., W. J. R., A. S.] and Medicinal Chemistry [M. B., J. B., H. N.], Boehringer Ingelheim Pharma KG, 88397 Biberach, Germany
Abstract
Pin1, a member of the parvulin family of peptidyl-prolyl cis-trans isomerases (PPIases) has been implicated in the G2-M transition of the mammalian cell cycle. Pin1 interacts with a series of mitotic phosphoproteins, including Polo-like kinase-1, Cdc25C, and Cdc27, and is thought to act as a phosphorylation-dependent PPIase for these target molecules. Pin1 recognizes phosphorylated serine-proline or threonine-proline peptide-bonds in test substrates up to 1300-fold better than in the respective unphosphorylated peptides. To test directly whether Pin1 regulates the G2-M transition and/or progression through mitosis by catalyzing phosphorylation-dependent prolyl isomerization of essential mitotic targets, we examined the consequences of Pin1 depletion, achieved by (a) overexpression of Pin1 antisense RNA, (b) overexpression of dominant-negative Pin1, and (c) by a known small-molecule Pin1-PPIase inhibitor, juglone. The results of all of the three lines of investigation show that the catalytic activity of Pin1 is essential for tumor cell survival and entry into mitosis.
Introduction
The cell cycle of eukaryotic cells is regulated by the coordinated
phosphorylation and dephosphorylation of proteins and their
ubiquitin-dependent proteolytic degradation. For example, entry into
mitosis requires
Cdc23
and Cyclin B, which assemble into a kinase complex responsible for the
phosphorylation of important mitotic proteins at Ser/Pro and Thr/Pro
motifs. Cyclin B itself must be degraded by the anaphase
promoting complex/cyclosome complex to allow exit from mitosis.
In Aspergillus nidulans, an additional kinase, NIMA, is
required for proper progression through mitosis (1)
. Cells
lacking functional NIMA arrest in G2, whereas
deregulated expression of NIMA induces premature mitosis
(2)
. A NIMA-like pathway has been proposed for vertebrate
cells as well (3)
. Recently, a protein interacting with
NIMA (Pin1) has been identified by a yeast-two-hybrid screen, using
NIMA as bait (4)
. Pin1 is a small
Mr 18,000 protein with two
distinct functional domains, namely a
NH2-terminal WW domain and a COOH-terminal
PPIase domain). Pin1 was found to be a homologue to the
Drosophila dodo protein (5)
, and it complements
the essential Saccharomyces cerevisiae homologue, Ess1/Ptf1
(4)
. Deregulation of Pin1-levels in yeast, human HeLa
cancer cells, and Xenopus laevis egg extracts has the
reciprocal phenotype to NIMA manipulation (4, 5)
, which
suggests that Pin1 negatively regulates entry into mitosis and is
essential for progression through mitosis. However, the deletion of the
Pin1 homologue in Drosophila melanogaster
(6)
and in the mouse (7)
did not result in
readily observable phenotypes. The 39-amino-acid WW domain of Pin1 acts
as a phosphoserine- and phosphothreonine-interaction module involved in
the binding of at least 30 mitotic phosphoproteins, including Cdc25C,
Plk1, Cdc27 (5, 8, 9)
and phosphorylated  protein
(10)
. Other functions for the Pin1-WW domain have been
suggested, including regulation of subcellular localization, regulation
of nuclear transport, transcription-promoting activity
(11)
, and pre-mRNA 3'-end formation (12)
. The
COOH-terminal domain of Pin1 is a phosphorylation-dependent PPIase that
catalyzes the cis-to-trans isomerization of
phosphoserine-proline and phosphothreonine-proline bonds with up to
1300-fold selectivity compared with unphosphorylated substrates
(13)
. Initial biochemical and crystal structure analysis
revealed that Cys-113, His-59, His-157, and Ser-154 form a catalytic
cascade with Cys-113 proposed to function as the general base in a
nucleophilic reaction, and Lys-63, Arg-68, and Arg-69 bind the
phosphate group of the substrate (13, 14)
. Pin1 may, thus,
act as a molecular switch by binding a subset of proteins
phosphorylated at Ser/Thr-Pro motifs and regulating their
biological activity through the isomerization of peptidyl-prolyl bonds
(14)
. However, proof that the PPIase activity of Pin1 is
required for its function in vertebrate cells is still lacking. In this
study, we addressed the importance of the Pin1 enzymatic activity in
human tumor cell-lines by transient overexpression of Pin1 dominant
negative proteins, by depletion of endogenous Pin1 through
overexpression of Pin1-antisense RNA, and through experiments with
the known Pin1-PPIase inhibitor, juglone.
protein
(10)
. Other functions for the Pin1-WW domain have been
suggested, including regulation of subcellular localization, regulation
of nuclear transport, transcription-promoting activity
(11)
, and pre-mRNA 3'-end formation (12)
. The
COOH-terminal domain of Pin1 is a phosphorylation-dependent PPIase that
catalyzes the cis-to-trans isomerization of
phosphoserine-proline and phosphothreonine-proline bonds with up to
1300-fold selectivity compared with unphosphorylated substrates
(13)
. Initial biochemical and crystal structure analysis
revealed that Cys-113, His-59, His-157, and Ser-154 form a catalytic
cascade with Cys-113 proposed to function as the general base in a
nucleophilic reaction, and Lys-63, Arg-68, and Arg-69 bind the
phosphate group of the substrate (13, 14)
. Pin1 may, thus,
act as a molecular switch by binding a subset of proteins
phosphorylated at Ser/Thr-Pro motifs and regulating their
biological activity through the isomerization of peptidyl-prolyl bonds
(14)
. However, proof that the PPIase activity of Pin1 is
required for its function in vertebrate cells is still lacking. In this
study, we addressed the importance of the Pin1 enzymatic activity in
human tumor cell-lines by transient overexpression of Pin1 dominant
negative proteins, by depletion of endogenous Pin1 through
overexpression of Pin1-antisense RNA, and through experiments with
the known Pin1-PPIase inhibitor, juglone.
Results
Catalytic Activity of Pin1 and Pin1 Mutant Proteins.
To study Pin1 at the biochemical and cellular level, wild-type Pin1 and
a set of mutant proteins were characterized in vitro and in
human tumor cell lines using transient transfection experiments. For
biochemical characterization, Pin1, the prolyl-isomerase domain
(PPIase), and a set of Pin1 mutants, including Pin1-C113A (C113A),
Pin1-R68L (R68L), Pin1-R69L (R69L), and the double point-mutant
Pin1-R68,69L (R68,69L) were expressed in Escherichia coli
and purified to homogeneity. The rate constant of prolyl isomerization
was determined by a protease-coupled isomerase assay (15)
.
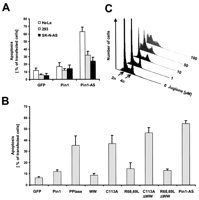
When increasing amounts of the various proteins were incubated with the
phosphorylated substrate, Ac-AApSPR-pNA, efficient isomerization was
observed in the presence of full-length Pin1 and the PPIase domain
(Fig. 1A![]() and Table 1
and Table 1![]() ). In contrast, Pin1-R68,69L exerted significant isomerase activity only
at protein levels >1000 ng, whereas C113A was catalytically inactive
in the protein range tested (Fig. 1A
). In contrast, Pin1-R68,69L exerted significant isomerase activity only
at protein levels >1000 ng, whereas C113A was catalytically inactive
in the protein range tested (Fig. 1A![]() ). Calculation of the
specificity constant,
kcat/Km,
revealed that Pin1 catalyzed cis-to-trans
isomerization approximately 400-fold more efficiently than R68,69L,
2-fold better than R68L, and 100-fold better than R69L, respectively
(Table 1)
). Calculation of the
specificity constant,
kcat/Km,
revealed that Pin1 catalyzed cis-to-trans
isomerization approximately 400-fold more efficiently than R68,69L,
2-fold better than R68L, and 100-fold better than R69L, respectively
(Table 1)![]() . When a nonphosphorylated substrate was used, Pin1 and
R68,69L were equally active at high enzyme concentrations, whereas
C113A again showed only little activity (Fig. 1B
. When a nonphosphorylated substrate was used, Pin1 and
R68,69L were equally active at high enzyme concentrations, whereas
C113A again showed only little activity (Fig. 1B![]() ; Table 1
; Table 1![]() ).
These data suggest that the side chain of C113 is required for the
catalytic reaction per se, whereas the two basic residues
R68 and R69 coordinate the binding of the phosphorylated substrate,
with R69 being more important than R68.
).
These data suggest that the side chain of C113 is required for the
catalytic reaction per se, whereas the two basic residues
R68 and R69 coordinate the binding of the phosphorylated substrate,
with R69 being more important than R68.
|
|
|
 WW), which suggests that these molecules
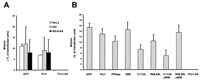
act as dominant-negative proteins. In contrast, cells transfected with
the WW domain, R68,69L, and R68,69L lacking the WW domain
(R68,69L-WW) showed no
significant increase in the number of apoptotic nuclei (Fig. 3B
WW), which suggests that these molecules
act as dominant-negative proteins. In contrast, cells transfected with
the WW domain, R68,69L, and R68,69L lacking the WW domain
(R68,69L-WW) showed no
significant increase in the number of apoptotic nuclei (Fig. 3B
|
 -tubulin, nuclear lamin, and phosphorylated histone H3 as
markers. As shown in Fig. 5A
-tubulin, nuclear lamin, and phosphorylated histone H3 as
markers. As shown in Fig. 5A
|
|
WW showed mitotic indices comparable with the GFP control
cells, whereas mitosis was significantly reduced in cells
overexpressing the PPIase domain, C113A, or C113A-WW
(P < 0.05, according to Student’s test). Consistent
with the experiments shown in Fig. 3C
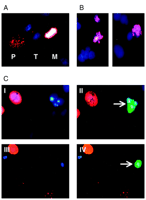
Impaired Pin1 Function Induces Apoptosis in Interphase.
To study in closer detail at which stage of the cell cycle Pin1
function is essential for cell survival, apoptotic cells that were
transfected with GFP and Pin1-antisense were inspected by
immunofluorescence for the presence of phosphorylated histone H3. As
shown in Fig. 5C![]() , phosphorylated histone H3 is not detected in apoptotic
nuclei of Pin1-depleted cells, which suggests that these cells do not
die in mitosis. In contrast, phosphorylated histone H3 is present in
cells dying in metaphase after treatment with Taxol (Fig. 5B
, phosphorylated histone H3 is not detected in apoptotic
nuclei of Pin1-depleted cells, which suggests that these cells do not
die in mitosis. In contrast, phosphorylated histone H3 is present in
cells dying in metaphase after treatment with Taxol (Fig. 5B![]() ).
).
Discussion
Depletion of Pin1 by overexpression of Pin1 antisense RNA induces
mitotic arrest and apoptosis in human tumor cells (4)
.
However, the Pin1 protein comprises distinct functional domains,
including the WW and PPIase domains, and whether the enzymatic activity
of the PPIase domain is required for this Pin1-mediated mechanism has
not been previously defined. To address this question, GFP-tagged
proteins—namely, Pin1, the isolated WW domain, the PPIase domain, and
a set of Pin1-mutants—impaired either in catalysis (C113A,
C113A-WW), or substrate recognition (R68,69L,
R68,69L-WW), were overexpressed in human tumor cell
lines.
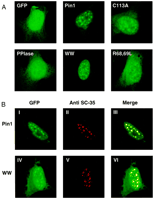
Transient transfection of HeLa cells with GFP-tagged Pin1 and the WW
domain resulted in a predominantly nuclear localization, with
high-lighting of nuclear speckles (Fig. 2)![]() . In contrast, the PPIase
domain showed no preferential nuclear staining but, instead, was
distributed throughout the cell. The mutants C113A and R68,69L exerted
a more diffuse but still speckled nuclear staining. These data suggest
that the WW domain is essential and sufficient for the retention of
Pin1 in nuclear speckles. However, additional interactions between
cellular substrates and the active site of Pin1 may either influence
the affinity toward proteins located in nuclear speckles or influence
the nuclear import of Pin1. The presence of Pin1 in nuclear speckles
may, therefore, reflect important WW domain as well as active-site
interactions. Functionally, nuclear speckles could serve as storage
sites for Pin1 as they do—it has been suggested—for splicing factors
(19)
.
. In contrast, the PPIase
domain showed no preferential nuclear staining but, instead, was
distributed throughout the cell. The mutants C113A and R68,69L exerted
a more diffuse but still speckled nuclear staining. These data suggest
that the WW domain is essential and sufficient for the retention of
Pin1 in nuclear speckles. However, additional interactions between
cellular substrates and the active site of Pin1 may either influence
the affinity toward proteins located in nuclear speckles or influence
the nuclear import of Pin1. The presence of Pin1 in nuclear speckles
may, therefore, reflect important WW domain as well as active-site
interactions. Functionally, nuclear speckles could serve as storage
sites for Pin1 as they do—it has been suggested—for splicing factors
(19)
.
Cells overexpressing the PPIase domain and the catalytic inactive
mutants C113A and C113A-WW showed a massive increase in the rate of
dead cells comparable with those observed in cells overexpressing Pin1
antisense RNA. All of the hallmarks of apoptosis were observed,
including chromatin condensation and fragmentation, membrane blebbing,
and disassembly of nuclear lamins (20)
. Presumably, the
mutants C113A and C113A-WW act as dominant negative versions of Pin1
by competing with endogenous Pin1 for the binding of physiological
substrates but are unable to catalyze the essential conformational
changes in substrate proteins required for cell survival. Our
data suggest that the localization of Pin1 in nuclear speckles is not
required for the induction of apoptosis because the overexpression of
the PPIase domain and C113A-WW, two mutants that do not bind to
these nuclear substructures, still induces cell death. Transfection of
Pin1, the WW domain, and the mutants R68,69L and
R69,69L-WW into human tumor cells did not result in an
increase of apoptosis, despite the fact that these proteins also were
at least 100-fold overexpressed compared with endogenous Pin1. These
data show that high levels of Pin1 and of the WW domain are tolerated
by cells and suggest that mutants like R68,69L and
R69,69L-WW do not compete with endogenous Pin1 for
essential substrates, presumably because of their reduced affinity
toward phosphorylated substrates.
An essential mitotic function for Pin1 has been proposed because
depletion of Pin1 resulted in abnormal mitotic phenotypes and
subsequent apoptosis (4)
. We reinvestigated the effects of
Pin1 depletion on mitosis using phosphorylation of histone H3 as a
mitotic marker. Depletion of Pin1 by antisense RNA resulted in a
complete absence of mitotic cells, in three of three human tumor cell
lines tested. Similarly, mitosis was reduced in cells overexpressing
the dominant negative proteins C113A and C113A-WW, which
suggests that the enzymatic activity of Pin1 is required for
G2-M transition.
In contrast to previous observations (4)
, we did not
observe abnormal mitotic events in cells overexpressing Pin1 antisense
RNA or the dominant negative mutants C113A and C113A-WW,
nor did we detect cells dying in mitosis. The morphological features
like cell rounding, chromatin condensation, lamin disassembly,
previously observed in Pin1depleted human tumor cells, actually
may reflect apoptosis in interphase rather than mitotic cell death
(20)
.
This findings were underscored by the use of juglone, the only known
inhibitor of the parvulin-like PPIases (16)
. In agreement
with the overexpression of C113A and C113A-WW and with previous
findings, juglone induced apoptosis (21)
and prevented
cells from entering into mitosis (22)
. However, because
juglone is a cysteine-reactive compound (16, 23)
and has
inhibitory activity on other proteins in vitro (23, 24)
, these findings need to be reevaluated with specific Pin1
inhibitors.
In agreement with our findings, the requirement of Pin1 in an interphase checkpoint has recently been described (25) . In Xenopus leavis egg extracts Pin1 participates in the replication checkpoint dependent on its catalytic activity. This novel function of Pin1 may explain why, in the absence of DNA damage, no apparent phenotypes were observed in mice lacking Pin1. (6)
In conclusion, our data show that the WW domain of Pin1 is required for the localization of Pin1 in nuclear speckles and suggest that phosphorylation-dependent peptidyl-prolyl cis trans isomerization catalyzed by Pin1 is essential for cell survival and for entry into mitosis. Consequently, these Pin1-PPIase-dependent mechanisms may serve as novel targets for the treatment of diseases associated with impaired apoptosis.
Materials and Methods
Plasmids.
Pin1 was cloned by reverse transcription PCR from K-562 cells using
primers 5'-CCGGATCCATGGCGGACGAGGAGAAGCTGC3' and
5'-CGAATTCAAGCTTCGAGGCCAGGCCTGGGCTCC-3'. Sitedirected mutations of
Pin1 were introduced using PCR-based techniques. As template, the Pin1
cDNA cloned into pCR2.1 (Invitrogen, Carlsbad, CA) was used. For
the generation of each mutant, two PCR reactions were carried out. For
C113A, the primer 5'-CAGCGACGCCAGCTCAGCCAAGGC-3'; for R68L, the primer
5'-CAGTCACTGCGGCCCTCGTCCTGG-3'; and for R69L, the primer
5'-CAGTCACGGCTGCCCTCGTCCTGG-3' were used together with the T7 primer.
For the second PCR, the corresponding complementary oligonucleotides
and the M13reverse primer were used. For each construct, the two
respective PCR products were annealed, a fill-in reaction was
performed, and the resulting DNA fragment was used for a third PCR with
T7 and M13reverse primers. The R68,69L mutation was generated by
introducing the R69L mutation in a construct already containing the
R68L mutation. The mutations were verified by automated sequencing. The
WW domain (amino acids 1–41) was generated by PCR using primers
5'-CCGGATCCATGGCGGACGAGGAGAAGCTGC-3' and
5'-AGCTAGCTGTTGCCGCTGGGCCGC-3'. The isomerase domain (amino acids
45–163) was generated using primers
5'-CCGGATCCATATGGGCAAAAACGGGCAGGGGGAGC-3' and
5'-CGAATTCAAGCTTCGAGGCCAGGCCTGGGCTCC-3'. For expression of GST-Pin1
fusion proteins in bacteria, the cDNAs encoding Pin1, the WW domain,
the PPIase domain and the various mutations were cloned into pGEX-4T-1
(Amersham Pharmacia Biotech, Freiburg, Germany). For mammalian
expression, the respective cDNAs were cloned in frame to GFP into
pEGFP-C1 (Clontech, Palo Alto, CA). The GFP-Pin antisense vector
was obtained by cloning the Pin1 PCR fragment with EcoRI,
BamHI into the EcoRI, BamHI sites of
pEGFP-C1 (Clontech).
Protein Expression and Purification.
Bacterial cells (XL1-blue; Stratagene, Heidelberg, Germany) that
harbored pGEX-4T-1-derived expression plasmids were grown in Luria
Bertani media. Protein expression was induced with 1
mM IPTG in midexponential growth phase. Four h after the
addition of IPTG, cells were harvested by centrifugation (10,000 x g, 10 min, 4°C) and the pellet was resuspended in lysis
buffer [20 mM disodium phosphate (pH 7.0), 1
mg/ml lysozyme, 10 mM EDTA, 1% (v/v) Triton
X-100, 10 µg/ml DNase, protease-inhibitors (Roche Diagnostics,
Mannheim, Germany)] at one-tenth of the culture volume. Cells were
lysed by three cycles of freeze-thawing, and unsoluble protein was
separated by centrifugation (10,000 x g, 10 min,
4°C). The supernatant was incubated with one-fifth volume of
preswollen glutathione Sepharose 4B (Amersham Pharmacia Biotech,
Freiburg, Germany) for 45 min at 4°C. Beads were separated from the
supernatant and washed by repeated centrifugation (300 x
g, 5 min, 4°C) and resuspension in PBS. Pin1 or
mutant proteins were released by incubation with bovine
-thrombin (1 µg/ml; Sigma, Deisenhofen, Germany) for 3 h at 30°C. Beads were separated from the supernatant by
centrifugation (300 x g, 10 min, 4°C). Bovine
-thrombin was removed by incubation of the supernatant with
benzamidine-Sepharose (Amersham Pharmacia Biotech, Freiburg, Germany)
at 4°C for 30 min and subsequent centrifugation (300 x
g, 10 min, 4°C). After addition of 1
mM DTT and 5 µg/ml Pefabloc (Serva, Heidelberg,
Germany) the supernatant was filtered (0.22 µm microcentrifuge filter
units; Millipore, Bedford) and concentrated (Centricon 10;
Amicon; Bedford, MA). Aliquots (20 µg/ml) were frozen in
liquid nitrogen and stored at -80°C.
Generation of Polyclonal Antisera against Pin1.
Two female rabbits were immunized with 12.5 µg of purified GST-Pin1
per animal, together with Friend’s adjuvans (Life Technologies, Inc.,
Karlsruhe, Germany). Prior to the s.c. (6 sides, 150 µl/side) and
muscle (4 sides, 25 µl/side) administration, the protein was adsorbed
to 2% AluGelS (Serva, Heidelberg, Germany). Immunization was repeated
three times with 25 µg of GST-Pin1, and animals were bled 2 weeks
after the final immunization.
Western Blot Analysis.
HeLa cell extracts were prepared by detaching the adherent cells with
trypsin/EDTA (Life Technologies, Inc., Karlsruhe, Germany), triplicate
washing with PBS, and lysis of the cell pellet by 4x SDS-loading
buffer (RotiLoad; Roth, Karlsruhe, Germany) at 95°C for 5 min. An
aliquot corresponding to 2 x 105 cells was
loaded on a 15% SDS-polyacrylamide gel. The proteins were separated by
electrophoresis and electroblotted onto nitrocellulose (Schleicher &
Schuell, Dassel, Germany). Endogenous or overexpressed Pin1 mutant
proteins were detected by incubation of the membrane for 2 h at
room temperature with rabbit anti-GST-Pin1 serum 1:250 in M-TBS
and subsequent incubation for 1 h with horseradish
peroxidase-linked donkey antirabbit serum (Amersham, Braunschweig,
Germany; 1:1000 in 5% M-TBS). After each step, membrane was washed
three times with 0.05% Tween 20 in TBS. Luminescent signals were
generated with SuperSignal West Pico substrate (Pierce, IL) and
recorded by autoradiography.
Determination of PPIase Activity.
The PPIase activity was measured as described (15)
.
Briefly, purified proteins (1–6000 ng) were preincubated with 72
µM substrate (Ac-AApSPR-pNA or Ac-AASPR-pNA; Interactiva,
Ulm, Germany) in a reaction volume of 60 µl in the presence of 35
mM HEPES (pH 7.8), 0.4 mg/ml BSA, and 2 mM DTT
for 30 min at 4°C. Proteolysis of substrate was initiated by the
addition of 70 µl trypsin (86 µg/ml) in 35 mM HEPES (pH
7.8). The release of p-nitroaniline was followed with a
microplate reader (MR5/7000/DIAS; Dynex Technologies, Denkendorf,
Germany) at 390 nm every 9 s. Resulting plots were fitted to a
first-order reaction and from the rate constant
kobs., the catalytic efficiency
kcat/Km
was calculated as described previously (15)
.
Cell Culture and Transfection of Mammalian Cells.
HeLa cells (ATCC, CCL-2) were grown in RPMI 1640 (Bio Whittaker, de
PetitRechain, Belgium), supplemented with 10% FCS; the human
kidney cell line 293 (ATCC, CRL-1573) and the neuroblastoma cell line
SK-N-AS (ATCC, CRL-2137) were grown in DMEM F-12 (Bio Whittaker)
supplemented with 10% FCS. Twenty-four h before transfection, 2 x 105 cells were seeded onto
poly-D-lysine-coated chamber slides and were incubated at
37°C in 5% CO2. Transfection was performed
using 1 µg of plasmid DNA and 5 µl of Fugene reagent (Roche
Diagnostics, Mannheim, Germany) according to the manufacturer’s
protocol.
Immunofluorescence Staining.
The following primary antibodies were used: antiphospho-histone-H3
rabbit serum (1:500 dilution; Upstate Biotechnology, Lake Placid,
NY); mouse monoclonal IgG recognizing spliceosome assembly
factor SC-35 (1:200 dilution, clone SC-35; Sigma, St. Louis,
MO); mouse monoclonal IgG PG-M3, directed against promyelocytic
leukemia (1:200 dilution; Santa Cruz Biotechnology, Santa Cruz,
CA); mouse monoclonal antilamin solution (undiluted; Progen,
Heidelberg, Germany); and mouse monoclonal anti--tubulin IgG (1:500
dilution; Sigma). Briefly, cells, fixed for 20 min with 2%
paraformaldehyde, were treated with 0.25% Triton X-100 for 10 min and
with 2% BSA/PBS for 30 min and were incubated for 1 h with 200
µl of the respective antibody-solution diluted in 2% BSA/PBS. For
fluorescent labeling, cells were further incubated with secondary
rhodamine-labeled goat antirabbit IgG, or rhodamine-labeled goat
antimouse IgG (1:1000 dilution; Jackson Immuno Research Laboratories,
West Grove, PA) in 2% BSA/PBS for 30 min. Cellular DNA was
stained with 2.5 µg/ml DAPI in PBS for 5 min. All of the steps were
performed at room temperature, and cells were washed two times with PBS
after each step. Finally, cells were mounted in 80% glycerol/PBS and
stored at 4°C. For documentation and microscopic analysis, a
fluorescence microscope (Axioplan 2; Zeiss, Jena, Germany) and a
laser scanning microscope (DM IRB/E; Leica, Bensheim, Germany)
were used.
Apoptosis.
The index of apoptotic cells was generally determined after DAPI
staining. Nuclei with highly condensed and fragmented DNA were scored
positive. This phenotype was confirmed by positive TUNEL staining of
the cells, which was performed according to the manufacturer’s
protocol (ApopTag fluorescein direct in situ apoptosis
detection kit; Intergen Company, Oxford, United Kingdom).
Fluorescence-activated Cell Sorting.
Ten thousand cells per analysis were used. Cells were incubated for
72 h with the indicated concentrations of juglone, fixed with 2%
paraformaldehyde for 20 min at room temperature, and permeabilized with
0.25% Triton X-100 in PBS by incubation for 5 min on ice. Cells were
pelleted by centrifugation (1000 x g, 5 min, 4°C),
resuspended in propidium iodine staining buffer (0.1% RNase, 10
µg/ml propidium iodine in PBS) and incubated for 20 min at room
temperature. The DNA content was analyzed using a FACS Calibur (Becton
Dickinson, Heidelberg, Germany).
Footnotes
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1 These two authors contributed equally to this
work. ![]()
2 To whom requests for reprints should be
addressed, at Department of Oncology Research, Boehringer Ingelheim
Pharma KG, Birkendorfer Strasse 65, 88397 Biberach an der Riss,
Germany. Phone: 49-7351-54-5240; Fax: 49-7351-54-5146; E-mail: andreas.schnapp{at}bc.boehringer-ingelheim.com ![]()
3 The abbreviations used are: Cdc, cell
division cycle; Ac, acetyl; DAPI, 4',6'-diamino-2-phenylindol; GFP,
green fluorescent protein; GST, glutathione
S-transferase; IPTG,
isopropyl-1-thio-ß-D-galactoside; NIMA, never in mitosis
A; pNA, para-nitroaniline; PPIase, peptidyl-prolyl
cis-trans isomerase; TUNEL, terminal
deoxynucleotidyl transferase-mediated nick end labeling; M-TBS, 5%
dried milk, 25 mM Tris (pH 7.4), 150 mM NaC. ![]()
Received for publication 12/15/99. Revision received 3/23/00. Accepted for publication 5/16/00.
References
protein. Nature (Lond.), 399: 784-788, 1999.[Medline]
 -crystallin inhibition by juglone. Biochem. Biophys. Res. Commun, 218: 648-652, 1996.[Medline]
-crystallin inhibition by juglone. Biochem. Biophys. Res. Commun, 218: 648-652, 1996.[Medline]
This article has been cited by other articles:
 |
A. S. Farrell, C. Pelz, X. Wang, C. J. Daniel, Z. Wang, Y. Su, M. Janghorban, X. Zhang, C. Morgan, S. Impey, et al. Pin1 Regulates the Dynamics of c-Myc DNA Binding To Facilitate Target Gene Regulation and Oncogenesis Mol. Cell. Biol., August 1, 2013; 33(15): 2930 - 2949. [Abstract] [Full Text] [PDF] |
||||
 |
G. Gudavicius, H. Soufari, S. Upadhyay, C. D. Mackereth, and C. J. Nelson Resolving the functions of peptidylprolyl isomerases: insights from the mutagenesis of the nuclear FKBP25 enzyme Biochm. Soc. Trans., June 1, 2013; 41(3): 761 - 768. [Abstract] [Full Text] [PDF] |
||||
 |
Y.-S. Lim, H. T. L. Tran, S.-J. Park, S.-A. Yim, and S. B. Hwang Peptidyl-Prolyl Isomerase Pin1 Is a Cellular Factor Required for Hepatitis C Virus Propagation J. Virol., September 1, 2011; 85(17): 8777 - 8788. [Abstract] [Full Text] [PDF] |
||||
 |
A. van der Horst and K. K. Khanna The Peptidyl-Prolyl Isomerase Pin1 Regulates Cytokinesis through Cep55 Cancer Res., August 15, 2009; 69(16): 6651 - 6659. [Abstract] [Full Text] [PDF] |
||||
 |
J. Kim, T. G. Choi, Y. Ding, Y. Kim, K. S. Ha, K. H. Lee, I. Kang, J. Ha, R. J. Kaufman, J. Lee, et al. Overexpressed cyclophilin B suppresses apoptosis associated with ROS and Ca2+ homeostasis after ER stress J. Cell Sci., November 1, 2008; 121(21): 3636 - 3648. [Abstract] [Full Text] [PDF] |
||||
 |
C. Fila, C. Metz, and P. van der Sluijs Juglone Inactivates Cysteine-rich Proteins Required for Progression through Mitosis J. Biol. Chem., August 1, 2008; 283(31): 21714 - 21724. [Abstract] [Full Text] [PDF] |
||||
|
|
F. Suizu, A. Ryo, G. Wulf, J. Lim, and K. P. Lu Pin1 Regulates Centrosome Duplication, and Its Overexpression Induces Centrosome Amplification, Chromosome Instability, and Oncogenesis Mol. Cell. Biol., February 15, 2006; 26(4): 1463 - 1479. [Abstract] [Full Text] [PDF] |
||||
|
|
S.-Y. Chen, G. Wulf, X. Z. Zhou, M. A. Rubin, K. P. Lu, and S. P. Balk Activation of {beta}-Catenin Signaling in Prostate Cancer by Peptidyl-Prolyl Isomerase Pin1-Mediated Abrogation of the Androgen Receptor-{beta}-Catenin Interaction Mol. Cell. Biol., February 1, 2006; 26(3): 929 - 939. [Abstract] [Full Text] [PDF] |
||||
|
|
F. Eckerdt, J. Yuan, K. Saxena, B. Martin, S. Kappel, C. Lindenau, A. Kramer, S. Naumann, S. Daum, G. Fischer, et al. Polo-like Kinase 1-mediated Phosphorylation Stabilizes Pin1 by Inhibiting Its Ubiquitination in Human Cells J. Biol. Chem., November 4, 2005; 280(44): 36575 - 36583. [Abstract] [Full Text] [PDF] |
||||
|
|
J. W. Hong, M. S. Ryu, and I. K. Lim Phosphorylation of Serine 147 of tis21/BTG2/pc3 by p-Erk1/2 Induces Pin-1 Binding in Cytoplasm and Cell Death J. Biol. Chem., June 3, 2005; 280(22): 21256 - 21263. [Abstract] [Full Text] [PDF] |
||||
|
|
J. D. Joseph, S. N. Daigle, and A. R. Means PINA Is Essential for Growth and Positively Influences NIMA Function in Aspergillus nidulans J. Biol. Chem., July 30, 2004; 279(31): 32373 - 32384. [Abstract] [Full Text] [PDF] |
||||
|
|
E. Bayer, S. Goettsch, J. W. Mueller, B. Griewel, E. Guiberman, L. M. Mayr, and P. Bayer Structural Analysis of the Mitotic Regulator hPin1 in Solution: INSIGHTS INTO DOMAIN ARCHITECTURE AND SUBSTRATE BINDING J. Biol. Chem., July 3, 2003; 278(28): 26183 - 26193. [Abstract] [Full Text] [PDF] |
||||
|
|
A. Ryo, Y.-C. Liou, K. P. Lu, and G. Wulf Prolyl isomerase Pin1: a catalyst for oncogenesis and a potential therapeutic target in cancer J. Cell Sci., March 1, 2003; 116(5): 773 - 783. [Abstract] [Full Text] [PDF] |
||||
 |
P. E. Shaw Peptidyl-prolyl isomerases: a new twist to transcription EMBO Rep., June 1, 2002; 3(6): 521 - 526. [Abstract] [Full Text] [PDF] |
||||
|
|
P.-J. Lu, X. Z. Zhou, Y.-C. Liou, J. P. Noel, and K. P. Lu Critical Role of WW Domain Phosphorylation in Regulating Phosphoserine Binding Activity and Pin1 Function J. Biol. Chem., January 25, 2002; 277(4): 2381 - 2384. [Abstract] [Full Text] [PDF] |
||||
 |
J.-L. Magnard, M. Yang, Y.-C. S. Chen, M. Leary, and S. McCormick The Arabidopsis Gene Tardy Asynchronous Meiosis Is Required for the Normal Pace and Synchrony of Cell Division during Male Meiosis Plant Physiology, November 1, 2001; 127(3): 1157 - 1166. [Abstract] [Full Text] [PDF] |
||||
|
|
H.-k. Huang, S. L. Forsburg, U. P. John, M. J. O'Connell, and T. Hunter Isolation and characterization of the Pin1/Ess1p homologue in Schizosaccharomyces pombe J. Cell Sci., October 15, 2001; 114(20): 3779 - 3788. [Abstract] [Full Text] [PDF] |
||||
 |
G. M. Wulf, A. Ryo, G. G. Wulf, S. W. Lee, T. Niu, V. Petkova, and K. P. Lu Pin1 is overexpressed in breast cancer and cooperates with Ras signaling in increasing the transcriptional activity of c-Jun towards cyclin D1 EMBO J., July 2, 2001; 20(13): 3459 - 3472. [Abstract] [Full Text] [PDF] |
||||
 |
T. Waerner, P. Gardellin, K. Pfizenmaier, A. Weith, and N. Kraut Human RERE Is Localized to Nuclear Promyelocytic Leukemia Oncogenic Domains and Enhances Apoptosis Cell Growth Differ., April 1, 2001; 12(4): 201 - 210. [Abstract] [Full Text] |
||||
|
|
I. Landrieu, B. Odaert, J.-M. Wieruszeski, H. Drobecq, P. Rousselot-Pailley, D. Inze, and G. Lippens p13SUC1 and the WW Domain of PIN1 Bind to the Same Phosphothreonine-Proline Epitope J. Biol. Chem., January 12, 2001; 276(2): 1434 - 1438. [Abstract] [Full Text] [PDF] |
||||
|
|
P.-J. Lu, X. Z. Zhou, Y.-C. Liou, J. P. Noel, and K. P. Lu Critical Role of WW Domain Phosphorylation in Regulating Phosphoserine Binding Activity and Pin1 Function J. Biol. Chem., January 25, 2002; 277(4): 2381 - 2384. [Abstract] [Full Text] [PDF] |
||||
|
|
I. Landrieu, B. Odaert, J.-M. Wieruszeski, H. Drobecq, P. Rousselot-Pailley, D. Inze, and G. Lippens p13SUC1 and the WW Domain of PIN1 Bind to the Same Phosphothreonine-Proline Epitope J. Biol. Chem., January 12, 2001; 276(2): 1434 - 1438. [Abstract] [Full Text] [PDF] |
||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HOME | HELP | FEEDBACK | SUBSCRIPTIONS | ARCHIVE | SEARCH | TABLE OF CONTENTS |
| Cancer Research | Clinical Cancer Research |
| Cancer Epidemiology Biomarkers & Prevention | Molecular Cancer Therapeutics |
| Molecular Cancer Research | Cell Growth & Differentiation |
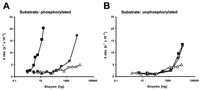
 , Pin1;
, Pin1;  , R68,69L;
, R68,69L;
 , C113A.
, C113A.